Factores de crecimiento y citocinas en la fisiopatología del cartílago Articular II
Introducción
El envejecimiento del cartílago articular se acompaña de una disminución del número de condrocitos por mm3 en mayor proporción en la zona superficial1,2 y de una disminución de la actividad de síntesis de las macromoléculas de la sustancia fundamental del cartílago en su totalidad.3
En los individuos de edad avanzada un número reducido de condrocitos podría asegurar la integridad anatómica del cartílago. Es conocido, además que las articulaciones de los individuos en la senescencia presentan signos frecuentes de alteraciones cartilaginosas4,5,6 y que la gravedad de estas lesiones es más notable en los sitios donde los ataques mecánicos son mayores; la degeneración del cartílago se acompaña de una necrosis de los condrocitos en la zona superficial, 17-9 y de una pérdida de proteoglicanos seguida de una ruptura de las fibras colágenas lo cual se traduce por la aparición de fisuras superficiales (Foto 1).
[su_spacer size=”30″]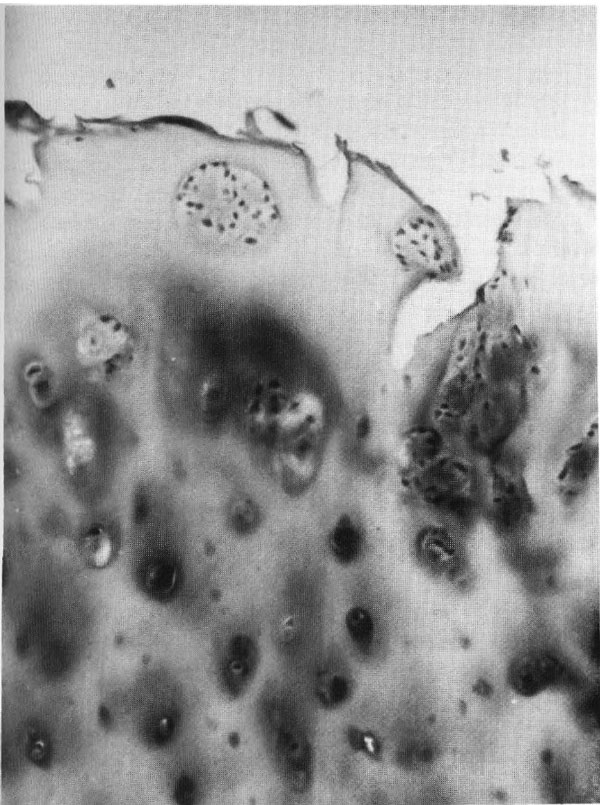
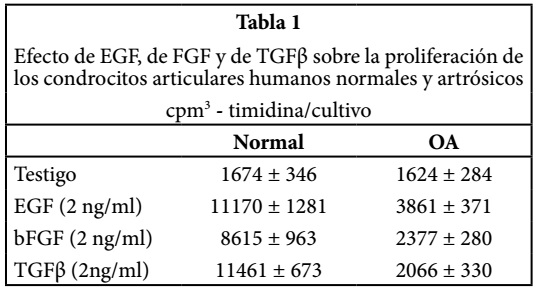
Foto 1. Cartílago artrósico tomado de la rótula de una mujer de 65 años.
La superficie del cartílago articular está lesionada y agrietada, los condrocitos de la capa superficial for man las clonas alrededor de las fisuras (flecha): la sustancia fundamenal(*) no tiene coloración la hemateina-eosina-safranina.
[su_spacer size=”30″]
Efectos de los Factores de Crecimiento y de las Citocinas sobre la Degradación de Proteoglicanos y del Colágeno
El efecto mayor de la IL-1 y del TNF-a sobre los condrocitos articulares es el de estimular la producción de las metaloproteasas, de las prostaglandinas y de los radicales libres, así como de acelerar el proceso catabólico en el cartílago.11-13 Los factores condrotropos, el TGF-β y el IGF-1 parecen bloquear este efecto14, 15 y el bFGF potencia este efecto de las citocinas;16 el mismo bFGF es también capaz de inducir un aumento de la producción de las proteasas por los condrocitos articulares.
A partir de nuestras investigaciones en los condrocitos articula res de ratas e TNFα parece inhibir la producción de la gelatinasa (MMP2) aún cuando la aparición de una gelatinasa de 92KDa no tiene efecto sobre la gelatinasa constitutiva de 62KDa También el GM-CSF, cuando actúa solo, no modifica los niveles de secreción de las gelatinasas, si no que potencia el efecto estimulador producido por IL-1 sobre la gelatinasa constitutiva de 62KDa. Por el contrario el GM-CSF, no actúa sobre el efecto inhibidor inducido por el TNFα.
Efecto de las Moléculas Extracelulares sobre la irregulación de la Degradación de Proteoglicanos y del Colágeno
Los fragmentos de ciertas moléculas de la sustancia fundamental del cartílago, tal como la fibronectina, tienen la propiedad de aumentar la síntesis de las metaloproteasas por los condrocitos.17 La fibronectina está largamente distribuida en el tejido conjuntivo, incluyendo el cartílago.18 Es una especie de pega a la que se adhieren las células que reconocen una secuencia de 3 aminoácidos arginina, glicina, y ácido aspártico, llamado también RGD.
La adhesión de las células se hace por intermedio de una proteína membranosa de la familia de las integrinas α5βI o VA5, presente en los condrocitos19 esta integrina se une a los filamentos de la actina del citoesqueleto cuya presencia determina la forma red onda o aplanada de la célula.
La adhesión de las células a las moléculas extracelulares produce una señal que incidirá sobre la actividad mitótica y metabólica. Sucesivamente, la incubación de fragmentos de cartílagos o los condrocitos aislados con el fragmento de 120KDa, que provienen de la fibronectina o con péptidos sintéticos que contienen la secuencia RGD de unión con las integrinas, conduce a un aumento de la degradación de proteoglicanos.17 En ese sistema, la fibronectina nativa no es activa.
La degradación de proteoglicanos se acompaña de una elevación de la estromelisina y de la colagenasa en el medio de cultivo20. Cuando los mismos productos son inyectados in vivo en la rodilla de rata éstos provocan un aumento de la densidad celular en el líquido sinovial, al igual que un aumento de los niveles de proteoglicanos y de las metaloproteasas.20
Dado que los niveles de los fragmentos de la fibronectina se elevan en el líquido sinovial de los enfermos afectados de artritis reumatoide o de artrosis,21,22 este mecanismo de regulación de la actividad de proteasas del cartílago puede en cierta medida contribuir a desencadenar los fenómenos observados.
Los Factores de Crecimiento y el Envejecimiento de los Condrocitos
Durante el envejecimiento muchas de las células del organismo, entre ellas los condrocitos, pierden progresivamente la capacidad de dividirse y responden débilmente a los factores de crecimiento.23 La causa por la cual se produce este fenómeno no es conocida; la pérdida de los receptores específicos y/o la disminución de su afinidad por el factor no ha sido encontrada en la mayoría de los casos. Sin embargo, este es el caso para ciertos factores, por ejemplo, el EGF Por el contrario, en otros, el número de receptores específicos por célula aumenta aún cuando la respuesta celular al factor disminuya, como ocurre con el factor IGF- 1.
En ciertos factores como la endotelina-1 (ET-1) los dos parámetros, la respuesta celular y la densidad de los receptores, aumentan paralelamente con la edad en su mayor parte, los receptores funcionan normal mente, la unión del factor con su receptor determina una señal que avanza en la célula hasta el núcleo donde parece encontrarse el sitio del bloqueo En efecto las células senescentes o envejecidas no son capaces de expresar ciertos factores necesarios para la activación de los genes que comandan la entrada de las células en la mitosis. Uno de los factores es el cfos24 que es un proto-oncógeno y puede aparecer inmediatamente después de una estimulación mitogénica y formar con otro proto-oncógeno cjun, el complejo cfos/cjun que se fija normalmente sobre el gen promotor que comanda la síntesis de ADN.25
Efectos de los Factores de Crecimiento de las Citocinas sobre el Condrocito Artrósico
La necrosis de los condrocitos de la zona superficial del cartílago articular es probablemente la lesión que desencadena la alteración del cartílago en la artrosis.1 La disminución progresiva de la densidad celular con la edad es un fenómeno universal y fisiológico que ocurre también en el cartílago articular.1, 2 La muerte celular fisiológica es conocida con el nombre de apoptosis. (Foto 2) Los aspectos reumatológicos de la apoptosis han sido descritos recientemente.26 En el curso del desarrollo de los órganos embrionarios o durante el funcionamiento de ciertos órganos y tejidos adultos como es el caso de la regeneración de la epidermis, la muerte celular es programada y desencadenada por la activación de ciertos genes,27 aunque también puede ser inducida por muchos agentes físicos o químicos pero en este caso se trataría de una muerte patológica, diferente a la apoptosis. Entre los factores capaces de inducir la apoptosis se encuentran la IL-1. TNFα, los glucocorticoides y la Hipoxia.26
[su_spacer size=”30″]
Foto 2. Condrocito necrótico (apoptosis) observado al microscopio electrónico en la zona profunda del cartílago articular artrósico. Los signos de apostosis están bien marcados: densificación y retracción del citoplasma y del núcleo, brotes de la membrana plasmática (flecha), alteración de las aorganelas citoplasmáticas. Coloración al acetato de uranilocitrato de plomo. Agrand.: 45.000X.
[su_spacer size=”30″]
Desde hace mucho tiempo hemos insistido en nuestro trabajo sobre el papel que juega la hipoxia del cartílago como factor responsable de la necrosis de los condrocitos en la artrosis.10 Esta hipoxia podría deberse a que el líquido intersticial, que es el que asegura los cambios del condrocito, queda retenido y esto ocasionaría la pérdida de elasticidad del cartílago envejecido y por supuesto contribuiría a elevar el choque mecánico.
La isquemia del cartílago articular en la zona de hiperpresión y la muerte celular contribuirían finalmente a la activación y liberación de las sustancias nocivas tales como las enzimas proteolíticas y los radicales libres tóxicos.10 La destrucción de la matriz extracelular traería como consecuencia el reblandecimiento del cartílago, que se fisura bajo la acción de los choques mecánicos.
La aparición de estas fisuras facilitaría la difusión de varios factores que se encuentran depositados en el mismo, tales como el bFGF y el TGFβ, produciendo una activación de los condrocitos viables con aumento de las mitosis28 de la actividad metabólica29, 30 así como una hipe producción de enzimas31 actualmente disponemos de informaciones parciales, aunque bien avanzadas, referidas a la sensibilidad de los condrocitos artrósicos, a los factores de crecimiento, a las citocinas y a otros agentes elásticos que intervienen en el metabolismo del cartílago.
También se conoce que los condrocitos artrósicos son in sensibles al IGF-1, aún cuando de la misma manera el número de receptores específicos32 y los niveles circulantes de este factor33 aumentan en los enfermos que padecen de artrosis. En compensación, la sensibilidad y la IL-I34 de los condrocitos artrósicos cultivados en el TGF-β estaría aumentada.
Según nuestra experiencia, en los casos en donde los condrocitos fueron aislados a partir de los fragmentos de cartílago en dos enfermos; uno de 59 años, con una necrosis isquémica idiopática de la cabeza femoral, en la que la superficie del cartílago era normal y otro de 60 años con una coxartrosis, cultivados los condrocitos en las mismas condiciones, los tapices de condrocitos artrósicos respondieron muy débil mente a los 3 factores de crecimiento estudiados (el EGF, bFGF y el TGF-β) (ver Tabla 1).